 咨詢熱線:0632-3279666
咨詢熱線:0632-3279666  城市分站
城市分站  聯(lián)系我們
聯(lián)系我們 

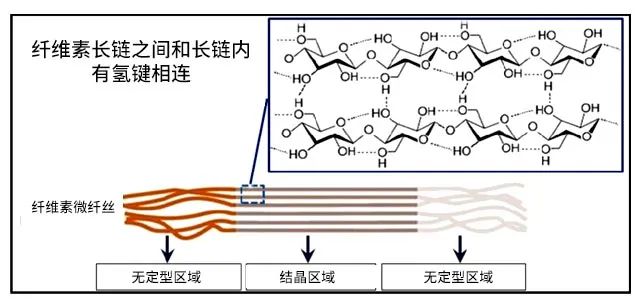
纖維素是由多個葡萄糖單元通過β-1,4鍵連接的多聚糖,分為無定形區(qū)和結(jié)晶區(qū)。在纖維素結(jié)晶區(qū)域,纖維素長鏈之間通過氫鍵緊密相連 (Nishyiama et al., 2002),長鏈內(nèi)部也有氫鍵相連接,排列緊密,β-1,4鍵被包裹在里面,這部分區(qū)域很難被酶解或者發(fā)酵(Ciolacu et al., 2011)。在纖維素?zé)o定型區(qū)域,長鏈之間沒有氫鍵連接。這部分纖維素相對比較疏松,酶有機會能夠接觸到里面的β-1,4鍵進行酶解。這也是酶解纖維素的主要區(qū)域。
纖維素的無定型區(qū)域與結(jié)晶區(qū)域示意圖
2纖維素分解酶Cellulolytic Enzyme
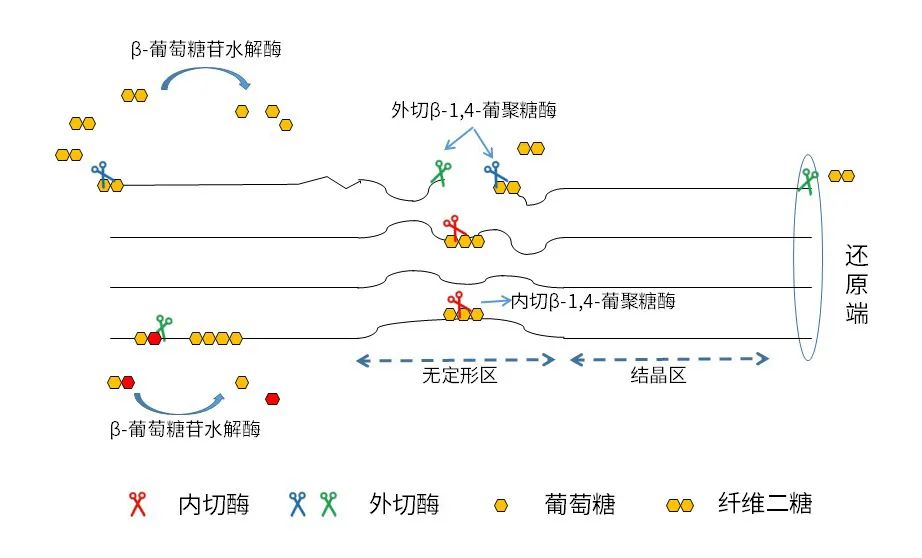
纖維素的降解區(qū)域主要在其無定形區(qū),分解無定型纖維素需要4種酶,分別是外切-葡聚糖酶、內(nèi)切-葡聚糖酶、纖維糊精酶和β-葡萄糖苷酶 (Mansfield et al., 1999; Zhang and Zhang, 2013)。外切-葡聚糖酶切割位于鏈兩端的糖苷鍵,內(nèi)切葡聚糖酶切割無定型纖維素長鏈的內(nèi)部糖苷鍵。β-葡萄糖苷水解酶和纖維糊精酶切割分解出來的短鏈鏈纖維素,生成單個葡萄糖 (Zorec, 2014)。小腸中完成的酶解,葡萄糖可以被小腸直接吸收利用,就像利用淀粉中的葡萄糖一樣,對能量的改善是顯著的。后腸發(fā)生的酶解,釋放的葡萄糖將被后腸微生物群利用,隨后合成短鏈脂肪酸,這些脂肪酸將被吸收和代謝為能量或脂質(zhì)沉積。
纖維素酶解和發(fā)酵的主要挑戰(zhàn)包括:(1)由于葡萄糖鏈之間的氫鍵結(jié)合,纖維素結(jié)晶部分的β-1,4糖苷鍵不可用;(2)豬后腸缺乏足夠的纖維素分解微生物;(3)纖維素酶解所需的β-葡聚糖酶活性較低,并且在后腸停留的時間短。
纖維素結(jié)晶區(qū)域的氫鍵需要通過化學(xué)處理才可以打開(Fahey等人,1993年),目前沒有酶能夠打開這些氫鍵。因此,纖維素的利用在很大程度上僅 限于其無定型部分。如果將這部分無定形纖維素充分降解,一方面可以貢獻額外的葡萄糖,另一方面,促進與之結(jié)合的其他營養(yǎng)物質(zhì)的充分利用。其中,纖維素酶是降解這一結(jié)構(gòu)的關(guān)鍵酶之一。
3希杰尤特爾纖維素酶CJYOUTELL Cellulase
纖維素酶與其他飼料酶一樣,這些酶要能夠抵抗胃部的低pH,并對胃蛋白酶和胰蛋白酶具有抵抗力。此外,由于在小腸中的轉(zhuǎn)運時間短,纖維素酶需要能夠在2至4小時內(nèi)作用,在消化液離開小腸之前釋放葡萄糖。
3.1希杰尤特爾纖維素酶酶學(xué)特性:
(1)自有專利菌種(US6190189B1)深層液體發(fā)酵。產(chǎn)品含有水解纖維素所需的三個酶:內(nèi)切β-1,4葡聚糖酶、外切β-1,4葡聚糖酶和β-葡萄糖苷酶。協(xié)同高x,酶解c底。
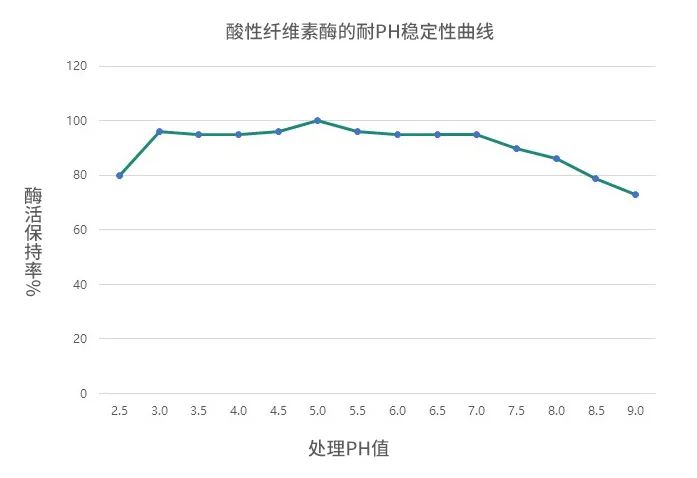
(2)希杰尤特爾纖維素酶耐溫耐酸性
耐pH范圍廣泛,在小腸內(nèi)保持高酶活,具有良好的耐溫性能,有效降解纖維素。
3.2希杰尤特爾纖維素酶應(yīng)用效果
3.2.1 對纖維結(jié)構(gòu)的破壞
電鏡觀察纖維形態(tài)(6000PFI),a為不添加纖維素酶,b為添加纖維素酶。纖維素結(jié)構(gòu)被破壞后,釋放出一些親水性物質(zhì),形成親水凝膠層。(Peng等, 2018)
3.2.2 對高纖維秸稈的水解
秸稈的主要成分是纖維素、半纖維素和木質(zhì)素,使用纖維素酶和木聚糖酶水解秸稈,觀察對纖維素的降解效果(慈元釗等,2019)
 郵箱:ren6669@126.com
郵箱:ren6669@126.com  電話:0632-3279666
電話:0632-3279666 
 地址:山東省棗莊市市中區(qū)孟莊鎮(zhèn)工業(yè)園區(qū)移山路6號
地址:山東省棗莊市市中區(qū)孟莊鎮(zhèn)工業(yè)園區(qū)移山路6號 
